Cours
Stabilité des allèles dans une population : Hardy-Weinberg




Les populations évoluent quand leur fréquence allélique change et donc que l’équilibre de Hardy Weinberg est modifié. Il existe 2 grands mécanismes évolutifs responsables du changement des fréquences alléliques : la sélection naturelle et la dérive génétique. Parfois l’évolution d’une population est telle qu’elle se coupe de la population d’origine et une nouvelle espèce apparaît : c’est la spéciation.
On définit la sélection naturelle comme le mécanisme qui fait que les individus d’une espèce présentant les caractères les plus avantageux pour survivre dans un environnement seront ceux qui vont plus facilement se reproduire. Ils vont peu à peu représenter la majorité de la population et donc modifier les caractères de l'espèce. Il existe plusieurs mécanismes de sélection naturelle.
La condition n° 5 du modèle de Hardy Weinberg est que pour que l’équilibre soit atteint, il ne doit pas y avoir de sélection zygotique : tous les zygotes formés lors de la fécondation c'est-à-dire les cellules-œufs à l’origine des individus, sont censés avoir la même espérance de vie pour le gène considéré.
Or dans le cas de certaines maladies autosomales récessives, les individus atteints n’atteignent pas l’âge de la reproduction, ne transmettant ainsi pas leurs allèles. C’est la sélection zygotique. Les fréquences mesurées dans les générations suivantes ne correspondent alors pas aux fréquences théoriques.
La mucoviscidose est une maladie rare qui touche principalement les voies respiratoires provoquant des infections à l’origine de toux et d’expectorations permanentes et invalidantes. Elle touche aussi le système digestif provoquant une obstruction des voies et des canaux digestifs à l’origine de troubles digestifs et hépatiques. Peu à peu les poumons se détruisent et l’insuffisance respiratoire s’aggrave.
Les individus atteints de mucoviscidose possèdent une protéine transmembranaire CFTR altérée dont le rôle en temps normal est d’excréter les ions chlorure à l’origine d’une fluidification du mucus des voies respiratoires et digestives. La mutation de cette protéine empêche l’excrétion des ions chlorures ce qui rend le mucus collant et visqueux.
L’espérance de vie des personnes atteintes de mucoviscidose était de 9 ans en 1960, de 25 ans pour celles nées dans les années 1990 et pour celles nées après 2000 elle peut atteindre 40 ans selon les traitements appliqués avec une médiane à 25 ans.
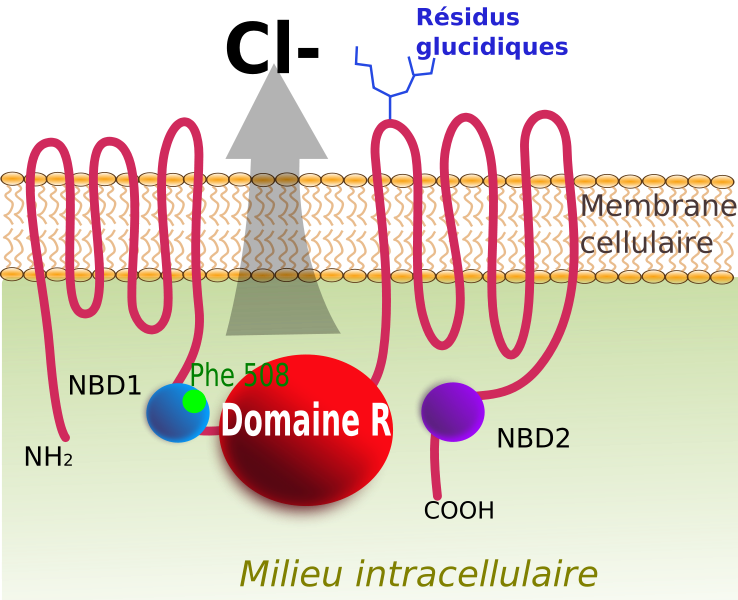
Protéine CFTR transmembranaire formant un canal à ions chlorure
Une étude épidémiologique génétique indique que sur une population de 54 900 français, 4 sont malades et 1 639 sont porteurs sains.
On notera l’allèle codant pour une protéine normale « CFTR » et l’allèle codant pour une protéine non fonctionnelle « cftr ». L’allèle « cftr » est récessif.
À partir des études épidémiologiques, on peut établir les fréquences mesurées dans la population :
u, la fréquence mesurée du génotype homozygote dominant (CFTR// CFTR), soit :
v, la fréquence mesurée du génotype hétérozygote (CFTR //cftr) soit :
w, la fréquence mesurée du génotype homozygote récessif (cftr // cftr) soit :
Calculons la fréquence attendue des génotypes dans la génération suivante. Pour cela procédons par étapes :
Calculons la fréquence des allèles dans la population étudiée.
Calculons les fréquences théoriques des génotypes de la descendance de cette population.
Probabilité d’apparition d’un homozygote dominant :
Probabilité d’apparition d’un hétérozygote :
Probabilité d’apparition d’un homozygote récessif :
Comparons les fréquences mesurées dans la population avec les fréquences théoriques attendues dans la descendance.
Pour le génotype homozygote dominant :
probabilité d’apparition d’un homozygote dominant :
Les 2 valeurs sont très proches.
Pour le génotype hétérozygote :
Probabilité d’apparition d’un hétérozygote :
Les 2 valeurs sont très proches.
Pour le génotype homozygote récessif :
Probabilité d’apparition d’un homozygote récessif :
La fréquence mesurée est nettement inférieure à la fréquence théorique attendue. Cela s’explique par la diminution de l’espérance de vie des individus atteints et ainsi une diminution de la transmission des allèles récessifs au sein de la population.
La condition n° 6 du modèle de Hardy Weinberg est que pour que l’équilibre soit atteint les individus doivent être pareillement viables et fertiles, c'est-à-dire qu’il n’y a, entre autres, pas de sélection d’un phénotype particulier.
Or quand la sélection du partenaire dépend d’un caractère sexuel variant, alors la fréquence de transmission des allèles est modifiée. Si certains mâles présentent un caractère peu attractif, ils se reproduiront moins et transmettront ainsi moins bien leurs allèles.
On peut donner comme exemple l’oiseau Euplecte à longue queue. Ce passereau commun en Afrique subit de grandes transformations en période de reproduction. De couleur beige striée de brun et de noir une grande partie de l’année, le mâle devient entièrement noir à l’exception d’une tâche rouge à l’épaule. Les plumes de sa queue tombent pour laisser place à des plumes ayant jusqu’à deux fois la longueur de son corps.
Cette longueur des plumes est contrôlée génétiquement. Il a été répertorié deux allèles conférant des tailles de plumes de queue différentes (longues ou courtes). Ce caractère s’il présente un handicap pour le vol comme chez le paon, est un avantage sélectif. En effet les femelles seront d’autant plus attirées par un mâle que les plumes de sa queue sont longues.

Euplecte en période de reproduction
Une étude a été réalisée sur trois populations d’euplectes comportant chacune 9 mâles répartis sur des territoires bien définis et non contigus. Ces mâles possèdent des plumes de queues de tailles identiques et la qualité des territoires mâles sont similaires.
Les ornithologues ont compté le nombre de nids actifs (ayant des œufs ou des petits). Quelle que soit la population, ils sont quasiment identiques comme le montre le document ci-dessous.
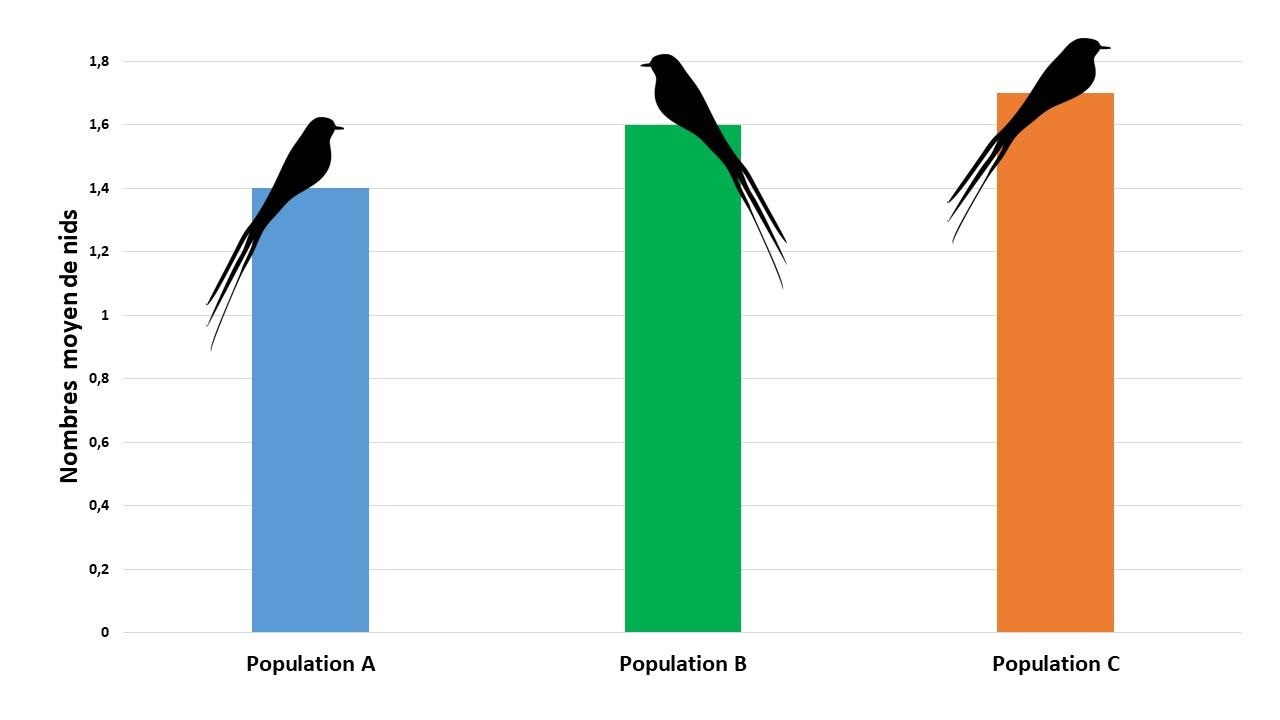
Graphique indiquant le nombre de nids actifs pour trois populations d’Euplectes
Pour vérifier le rôle de la longueur des plumes dans l’attraction sexuelle, les ornithologues ont sectionné les plumes des queues des mâles de la population A et les ont collées aux plumes des queues des mâles de la population C. Ils ont laissé la période de reproduction s’effectuer et ont comptabilisé les nids actifs.
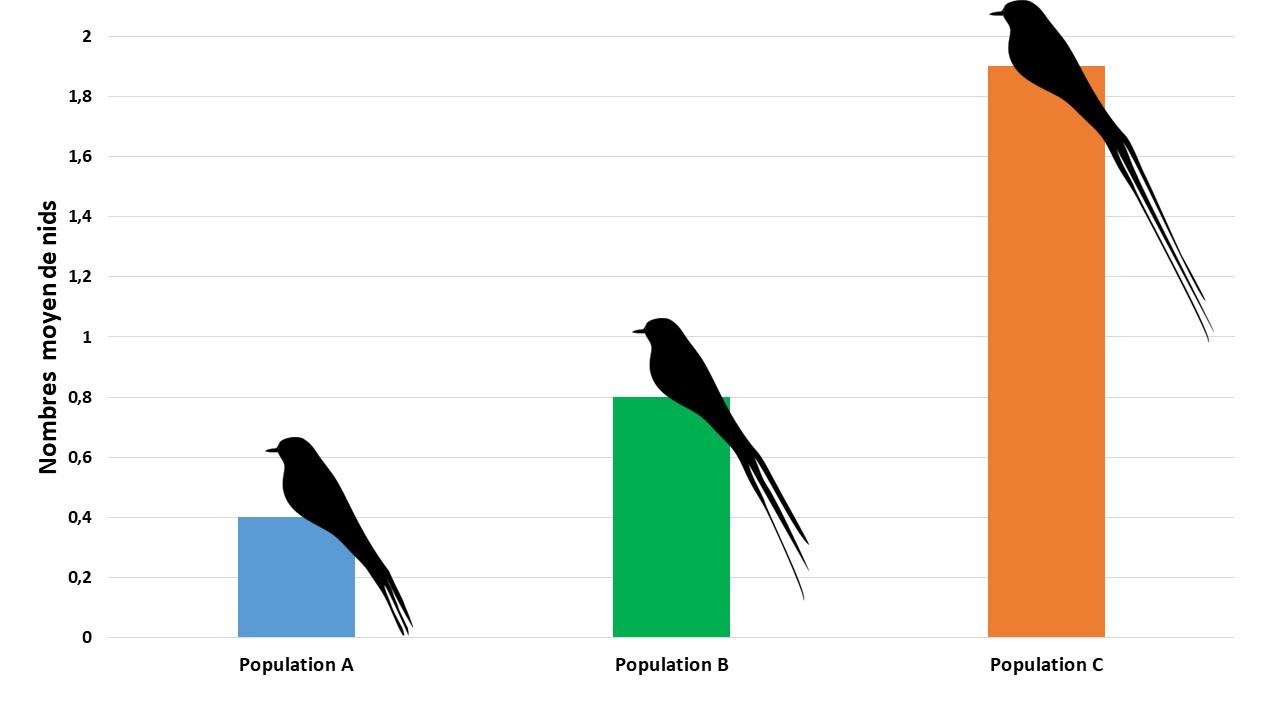
Graphique indiquant le nombre de nids actifs pour trois populations d’Euplectes après avoir rallongé artificiellement les plumes des queues des mâles de la population C
La population ayant les mâles possédant les plumes de queue rallongées artificiellement présentent un nombre de nids actif plus important que celle dont les mâles ont les plumes raccourcies.
Ainsi, si on revient à l’étude génétique de la population d’origine, concernant les allèles responsables de la longueur des plumes de la queue, on peut donc dire que dans toute population, en raison de l’attraction des femelles pour les mâles ayant les plumes de queue les plus longues, l’allèle « longue queue » se transmettra plus facilement que l’allèle « courte queue », modifiant ainsi sa fréquence au fur et à mesure des générations.
La condition n° 3 dit que pour que l’équilibre de Hardy Weinberg soit atteint il ne doit pas y avoir de mutation dans la population car celle-ci modifie un caractère perturbant ainsi la relation existante entre l’être vivant et son biotope. Peuvent alors s’en suivre une sélection des individus et donc une perturbation de l’équilibre.
On peut citer le cas des phalènes du bouleau. La Phalène du Bouleau, Biston betularia, est un papillon de nuit de la famille des Geometridae, commun en Europe du Nord. Ces papillons de nuit passent la journée posés sur les troncs des bouleaux, où ils s’abritent des prédateurs (principalement des oiseaux) car ils se fondent sur les lichens gris poussant que les troncs de couleur pâle de ces arbres. Cette espèce présente un phénotype alternatif : on distingue une forme claire (typica) qui est la forme originelle et une forme sombre (carbonaria) observée pour la première fois à Manchester en Angleterre en 1848.

Biston betularia, forme « typica »

Biston betularia, forme « carbonaria »
Du point de vue génétique, la couleur est liée à un gène qui existe sous deux formes alléliques :
L’allèle c récessif qui est l’allèle responsable de la coloration typica (claire).
L’allèle C dominant qui est responsable de la coloration carbonaria (noire).

Couple de Biston betularia

Phalène du bouleau de type carbonaria et de type typica sur un tronc de bouleau
Au milieu du XIXème siècle, avant la révolution industrielle, la forme typica était largement majoritaire dans les populations anglaises. En temps normal l’allèle C est défavorable à l’espèce car il rend les papillons foncés et bien visibles sur les lichens gris clair des troncs de bouleaux n'échappant donc pas aux prédateurs. Ainsi au sein de la population peu de papillons noirs peuvent se reproduire. En temps normal, l’allèle c est favorable à l’espèce car il permet aux papillons (c//c) d’être de couleur claire et de se dissimuler sur les lichens. Ils peuvent survivre, se reproduire et transmettre cet allèle à leur descendance.
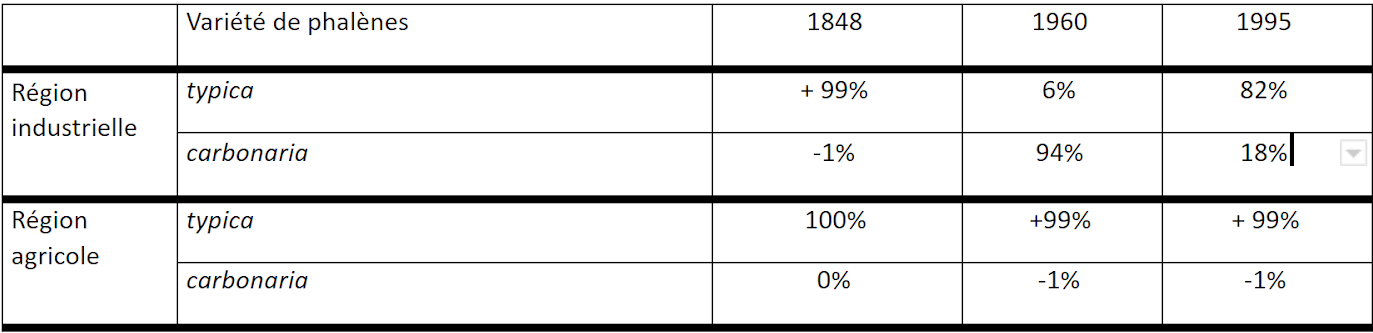
Évolution des fréquences de phalènes
Apparue suite à une mutation, la fréquence de la forme carbonaria s'est accrue rapidement et les individus mélaniques ont ensuite été observés dans les autres régions industrielles de l'Angleterre. En effet, après 1850, avec la révolution industrielle, l’augmentation de la population humaine en ville et l’usage généralisé du charbon, les particules de fumée augmentèrent spectaculairement à Londres et dans ses environs, dont la ville industrielle de Manchester, créant un brouillard dense appelé « smog ».
Les troncs des bouleaux, sur lesquels les phalènes se tenaient, commencèrent lentement à perdre leurs lichens et à noircir en se recouvrant de suie et de poussière de charbon. Ainsi seuls les papillons de couleur foncée ont pu se camoufler, survivre et donc se reproduire et transmettre l’allèle C à leur descendance. Au sein de la population peu de papillons blancs ont survécu et ont pu se reproduire : cette forme claire est donc devenue de moins en moins nombreuse. L'allèle C est donc devenu de plus en plus présent dans la population au fil des générations.
En 1950, la forme mélanique représentait 90% de la population dans cette région. Ce phénomène « apparition d’individus très sombres dans la deuxième moitié du 19ème siècle et l’augmentation rapide de leur fréquence » fut observée chez de nombreuses autres espèces de papillons de nuit, ainsi que chez différentes espèces d’insectes, d’araignées, et même d’oiseaux, pas seulement dans la région de Manchester, mais aussi dans les environs d’autres villes industrielles telles que Birmingham et Liverpool. On parla alors de "mélanisme industriel".
Si les régions très industrielles ont souffert dès le XIX ème siècle d’une importante pollution, les régions très agricoles du sud-ouest de l’Angleterre ont peu souffert de la pollution liée à la révolution industrielle du XIXème siècle. Dans ces régions, la forme carbonaria n’apparaît qu’à partir de 1932. À partir de 1950, l’Angleterre cherche à lutter contre la pollution liée aux industries, cela a eu pour effet de diminuer rapidement les émissions polluantes. Très rapidement les lichens se développèrent à nouveau sur les troncs d’arbres et on constate de nouveau une prédominance de l’allèle c.

Colonne Nelson pendant le grand smog de 1952
Une population est un ensemble d’individus de la même espèce vivant à proximité les uns des autres, se reproduisant plus fréquemment entre eux qu’avec ceux d’autres populations.
L’effectif de la population étant fini, l’équilibre de Hardy Weinberg ne peut être respecté car l’échantillonnage aléatoire des gamètes lors de la fécondation modifie les fréquences des allèles d’une génération à une autre : c’est la dérive génétique.
Cette théorie a été établie par Motoo Kimura en 1968.
La dérive génétique correspond donc à une modification de la fréquence d'un allèle et donc d’un génotype au sein d'une population, indépendamment des mutations ou de la sélection naturelle.
Dans toute population, il est statistiquement inévitable que certains allèles ne soient transmis par aucun adulte à sa descendance tout comme certains allèles ne soient pas transmis car certains individus n'ont pas de descendance du tout. Le nombre des allèles, c'est-à-dire la diversité allélique ou variabilité génétique, se réduit donc. Parmi les allèles « survivants », certains vont voir leur fréquence originelle diminuer ou au contraire augmenter.
La dérive génétique concerne tous les allèles même si l'impact sur les allèles neutres (c’est-à-dire qui ne confèrent ni avantages ni inconvénients) est plus important. Malgré tout un allèle favorable peut disparaître ou un allèle défavorable se fixer dans une population par dérive, ce qui est fréquent pour des populations aux tailles très réduites. Les effets de la dérive génétique sont d'autant plus importants que la population est petite, car les écarts observés par rapport aux fréquences alléliques y seront d'autant plus perceptibles.
Couplée à la sélection naturelle, elle est un des moteurs de l’évolution des populations.
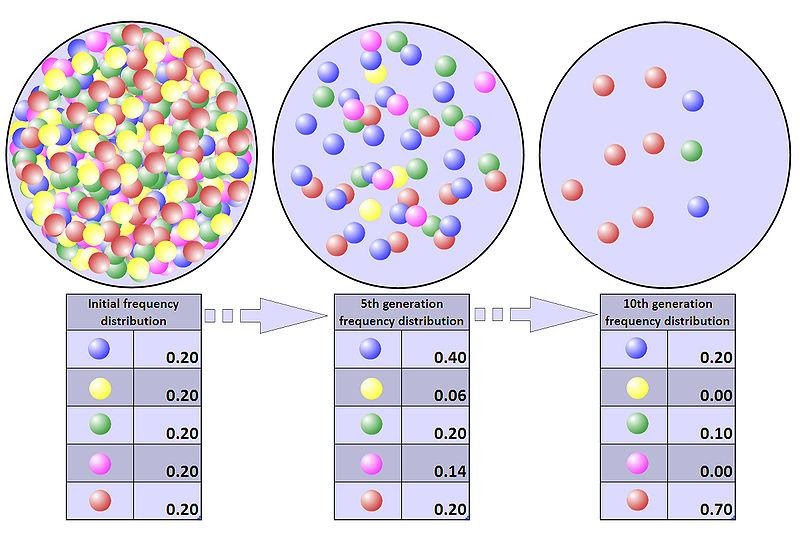
Représentation schématique de la dérive génétique couplée à la sélection naturelle
Au fur et à mesure de l’avancée des générations, seuls les individus ayant pu survivre se sont reproduits ce qui modifie la proportion des allèles (billes de couleur) dans la population.
La dérive génétique peut être provoquée par une baisse brutale du nombre d’individus dans une population, baisse qui est à l’origine d’une diminution de la diversité génétique. L’origine en est un évènement décimateur.
Aléa météorologique, épidémie ou encore une météorite comme il y a 65 Ma qui a provoqué la disparition des dinosaures et permis le développement des mammifères.
L’effet fondateur est également responsable de la dérive génétique.
Lorsqu'un nombre réduit d'individus se sépare d'une population plus vaste, pour aller coloniser une île ou un nouveau milieu, ces individus ne vont « emporter » qu'un échantillon d'allèles du pool d'allèles de la population mère, et ce, de manière que l'on suppose aléatoire. La nouvelle population peut donc présenter des fréquences génotypiques fort différentes de la population initiale. Cet écart peut changer radicalement le profil (allélique, génotypique et phénotypique) de la population fondatrice, par rapport à la population initiale. C’est une dérive génétique.
Du fait de la transmission aléatoire des allèles, on observe alors soit une sur-représentativité soit une sous-représentativité de certains allèles. Certaines mutations peuvent ainsi être conservées.
L'effet fondateur peut, s'il conduit à un isolement reproducteur, c'est-à-dire à la cessation d'échange de versions géniques entre l'ensemble des individus de l'espèce, aboutir à une spéciation, c'est-à-dire l’apparition d’une nouvelle espèce.
Les lions du cratère Ngorongoro (situé en Afrique de l’Est, Tanzanie) forment une petite population d’une centaine d’individus et sont issus de la grande population voisine du parc naturel de Serengeti (d’environ 2 000 individus). Les 2 milieux de vie sont très semblables mais séparés par une barrière géographique naturelle, les remparts du cratère.
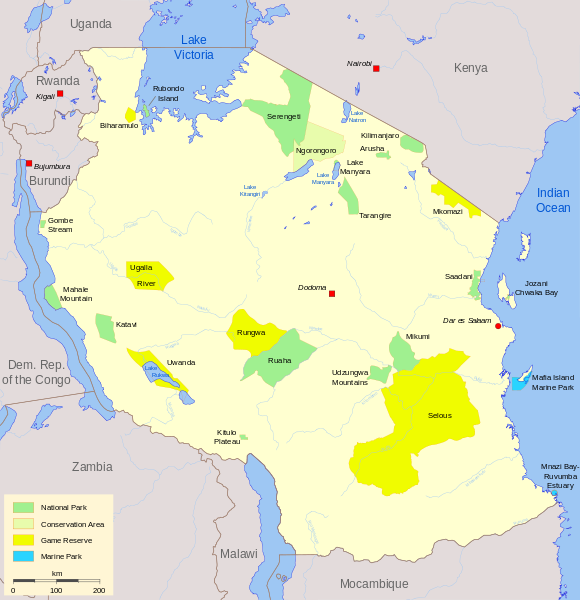
Localisation des parcs du Serengeti et du cratère Ngorongoro
Parks Tanzania.svg par Bamse via wikimédia commons, CC-BY-SA-3.0-migrated https://commons.wikimedia.org/wiki/File:Parks_Tanzania.svg

Lions dans le cratère Ngorongoro

Lion du cratère Ngorongoro
Carte topographique du cratère Ngorongoro

Lion du parc du Serengeti
En 1962, une grave infection fit chuter brutalement l’effectif de la population des lions du cratère. Une dizaine de lions seulement survécurent. Ces lions survivants se sont reproduits exclusivement entre eux et la population retrouve actuellement une taille d’environ 100 individus. Dans les années 1990, des chercheurs ont étudié la diversité génétique entre les deux populations de lions. Ils ont mesuré la fréquence allélique de 4 gènes pris dans les deux populations de lions.
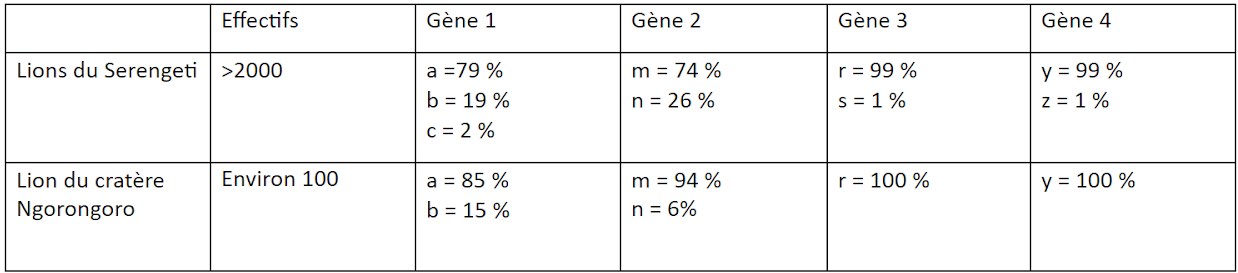
On observe que les gènes 1, 2, 3 et 4 existent dans les 2 populations de lions étudiées. Toutefois, le nombre et la fréquence de chaque allèle de chaque gène varie.
Il y a une variation du nombre d’allèles par gène : au sein de la population de lions du Serengeti, le gène 1 possède 3 allèles (a, b, c), alors qu’il n’existe qu’en 2 versions dans la population de lions Ngorongoro (a et b). De même, pour les gènes 3 et 4, les lions de Ngorongoro ne possèdent qu’un allèle (respectivement r et y), alors que les gènes 3 et 4 possèdent 2 allèles chacun dans la population de Serengeti.
Il y a également une variation de la fréquence des allèles : regardons le gène 2, qui existe sous 2 versions (allèles m et n) dans les 2 populations de lions. On remarque que la fréquence de ces allèles est différente : l’allèle m est très majoritaire (94%) par rapport à l’allèle n (6%), alors qu’au sein de la population de lions de Serengeti, les fréquences de ces allèles sont de respectivement 74% et 26%. Autrement dit, la population de lions Ngorongoto est moins diversifiée génétiquement que la population de lions Serengeti.
Ainsi la baisse de la diversité génétique observée dans la population de lions Ngorongoro s’explique de 2 façons :
L’infection de 1962 a fait très nettement chuter l’effectif de la population, donc certains allèles ont dû disparaître.
L’effectif réduit de cette population a amplifié le mécanisme de dérive génétique, qui baisse la diversité allélique au sein de la population de lions Ngorongoro, et accentue les différences génétiques entre les 2 populations de lions.
Avant la théorie de Darwin, une espèce était considérée comme permanente et stable et se définissait par rapport à un individu « type » : tous les individus qui lui ressemblent et qui sont interféconds sont rattachés à cette espèce, toute variations étant considérée comme une anomalie. Dans ce cadre, la biodiversité est discontinue, les groupes n’ayant aucun lien de parenté entre eux, et la question de l’origine et du devenir des espèces ne se pose pas et relève de la théologie.
Au XIX° siècle, la pensée évolutionniste modifie le concept d’espèce. La variabilité devient un caractère essentiel au moteur de l’évolution : la filiation entre les espèces s’impose. Dans l’arbre du vivant, une espèce est définie comme une sous-partie du réseau généalogique, un rameau qui diverge définitivement de la branche dont il est issu (c’est la spéciation).
Les critères permettant de définir une espèce sont les suivants :
Les critères phénétiques ou ressemblances morphologiques mais ils sont insuffisants en raison de la variabilité individuelle, par exemple le dimorphisme sexuel.
Les critères biologiques liés à l’interfécondité proposés en 1942 par Ernst Mayr « une espèce est une population ou en ensemble de populations dont les individus peuvent effectivement ou potentiellement se reproduire entre eux et engendrer une descendance viable et féconde, dans des conditions naturelles » ; ce critère est difficile à utiliser en pratique.
Critères génétiques liés à l’interfécondité : l’analyse ADN révèle des flux de gènes entre les populations.
Critères écologiques liés à l’interfécondité : exemple chez les végétaux, deux populations ayant des périodes de floraison différentes ou occupant des milieux de vie très différents ne peuvent se reproduire ensemble et donc n’appartiennent pas à la même espèce
La spéciation est, en biologie, le processus évolutif par lequel de nouvelles espèces se forment. Les espèces s'individualisent à partir de populations appartenant à une espèce d'origine.
La spéciation résulte de la dérive génétique et de la sélection naturelle, qui sont les deux moteurs de l'évolution.
Une espèce n'apparaît pas instantanément par une mutation conduisant à l'apparition d'un individu d'un type nouveau. Si un gène est modifié, le caractère contrôlé par ce gène se trouve également modifié. Les modifications peuvent soit présenter un avantage (capacité de survie, de reproduction), soit présenter un désavantage, soit être neutres. Cela dépend de la modification mais également de l'environnement (une même modification est un avantage quelque part et un désavantage ailleurs).
Les individus d’une espèce présentant les caractères les plus avantageux pour survivre dans un environnement seront ceux qui vont plus facilement se reproduire. Ils vont peu à peu représenter la majorité de la population et donc modifier les caractères de l'espèce.

Charles Darwin fut le premier à décrire le rôle de la sélection naturelle comme facteur de spéciation, dans son livre L'Origine des espèces (1859).
Une espèce peut donc être définie comme une population d’individus suffisamment isolés génétiquement des autres populations.
Elle n’existe cependant que sur une durée de temps finie.
Si un nouvel ensemble s’individualise au sein de la population, il y a isolement reproductif et spéciation (apparition d’une nouvelle espèce).
On nomme mécanisme d'isolement reproductif tout mécanisme empêchant ou limitant fortement l'hybridation de deux populations habitant la même région, même lorsqu'elles sont étroitement apparentées.
Si un ensemble d’individus se retrouve isolé géographiquement de la population principale, il y a isolement géographique.
Il y a extinction d’une espèce si l’ensemble des individus concernés disparaît ou alors si elle cesse d’être isolée génétiquement.
On distingue quatre modes de spéciations dans la nature, suivant le degré d'isolement d'une sous population par rapport aux autres :
Allopatrique (formation d'une barrière physique),
Péripatrique (par migration),
Parapatrique (par colonisation d'une niche écologique voisine),
Sympatrique (par colonisation d'une niche écologique ou comportementale dans la même aire de répartition).
On peut également créer des spéciations artificielles, par élevage, ou par des expériences de laboratoire.
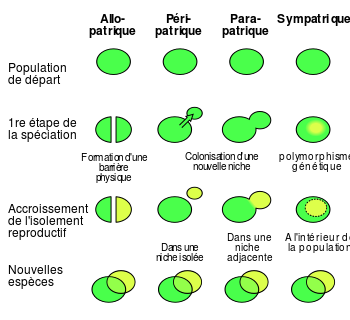
Modes de spéciation fr.svg, auteur wiki anglais, via wikimedia commons, CC-BY-SA-3.0-migré-avec-avertissements, https://commons.wikimedia.org/wiki/File:Speciation_modes_fr.svg
La spéciation allopatrique
L'effet fondateur est donc un des facteurs de la spéciation allopatrique (avec la sélection naturelle). L’apparition d’une barrière géographique est également un facteur de spéciation allopatrique.
On peut donner comme exemple l’apparition de 2 sous espèces de papillons du genre Zerynthia.
Les deux papillons Zerynthia cassandra et Zerynthia polyxena sont très semblables morphologiquement mais se différencient par la forme de leurs organes reproducteurs empêchant toute reproduction entre eux. Il s’agit donc d’espèces différentes dont les grandes ressemblances ont permis aux scientifiques de supposer qu’elles avaient une origine commune. Selon eux, elles proviennent d’une espèce ancestrale commune (Z) qui aurait subi des mécanismes aboutissant à la création de ces deux nouvelles espèces.
Pour valider cette hypothèse, ils ont étudié l’histoire géographique des aires où se trouvent ces deux espèces.

Zerynthia polyxena

Zerynthia cassandra

Dessin des appareils reproducteurs
Le quaternaire est la période géologique qui couvre les 2,5 derniers millions d’années (Ma). Cette période est caractérisée par des fluctuations climatiques. Celles-ci s’organisent en cycles comprenant chacun une phase glaciaire et une phase interglaciaire. La dernière phase glaciaire s’est achevée il y a 12 000 ans environ et a duré 100 000 ans. Chaque refroidissement climatique a provoqué l’extension des calottes polaires et des glaciers et durant les périodes de réchauffement interglaciaire, la calotte glaciaire est remontée vers le Nord.
Les glaciations entraînent une modification des aires de répartition des espèces végétales et animales. Ainsi pendant ces périodes de glaciation, les zones occupées par des forêts tempérées ont laissé la place à des steppes. Dans certains cas, certaines espèces adaptées au climat tempéré sont parvenues à se maintenir dans des zones plus au sud bénéficiant d’un climat relativement doux compatible avec leurs exigences : ces zones sont qualifiées de zones refuges.
Ainsi l’espèce Z a trouvé refuge dans les péninsules italiennes et grecques. Séparées par la mer Adriatique, les deux populations ont évolué indépendamment l’une de l’autre donnant naissance à 2 nouvelles espèces : Cassandra en Italie et Polyxena en Grèce. Depuis la fin de la glaciation, les populations sont remontées vers le nord et occupent des territoires contigus mais les différences de morphologie de l’appareil reproducteur empêchent toute hybridation.
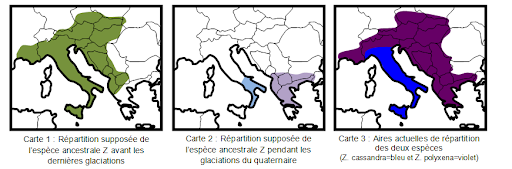
Aires de répartition des espèces Zerynthia
La spéciation sympatrique
Deux espèces de cichlidés peuplent le lac Victoria.
Ces deux espèces diffèrent par quelques caractères, notamment la couleur. L’absence d’hybrides dans le lac montre que ces 2 espèces ne se reproduisent pas entre elles. Le rayonnement solaire est absorbé par l’eau, mais cette absorption n’est pas égale pour toutes les composantes de la lumière. Dans les eaux des lacs, dans les couches d’eau supérieures, la sensibilité à la lumière bleue et à la coloration bleue est favorisée. À l’inverse, dans les couches d’eau profondes, c’est la sensibilité à la lumière rouge et à la coloration rouge qui prédomine.
Les poissons cichlidés du lac Victoria mesurent jusqu’à 15 cm de long. Les mâles de certaines espèces sont rouges (Pundamilia nyererei) et vivent jusqu’à -6 mètres de profondeur, les autres bleus (Pundamilia pundamilia) vivent entre 0 et -3 mètres de profondeur. Les femelles des deux espèces sont brunes et se ressemblent. Pour ces deux espèces, chaque mâle défend un territoire, visité par les femelles, qui choisissent leur mâle.
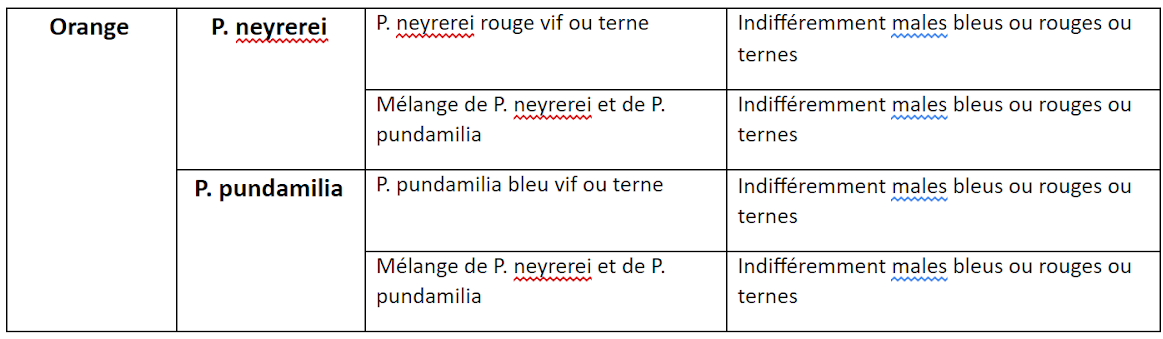
Pour comprendre comment se faisait le choix du partenaire, des expériences ont été réalisées en laboratoire. Des femelles de l’espèce P. nyererei ou P. pundamilia ont été placées dans un aquarium avec des mâles appartenant aux deux espèces. Cet aquarium a été éclairé soit par lumière naturelle soit par lumière orange qui empêche toute distinction des couleurs.

Pundamilia nyererei

Pundamilia pundamilia
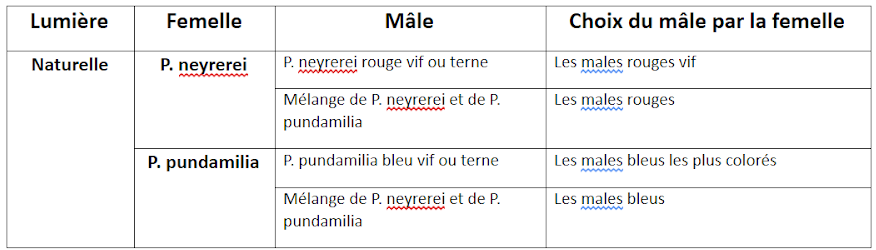
Résultats des expériences d’accouplement
Des études ont montré que chaque espèce de Cichlidés comporte un gène codant pour différentes versions de la protéine constitutive des photorécepteurs de la rétine. Ainsi, chaque espèce a des photorécepteurs ayant un spectre d’absorption différent.
Ainsi, des poissons placés dans un environnement dans lequel leur vision n’est pas adaptée auront du mal à trouver de la nourriture ou encore à trouver un partenaire sexuel.
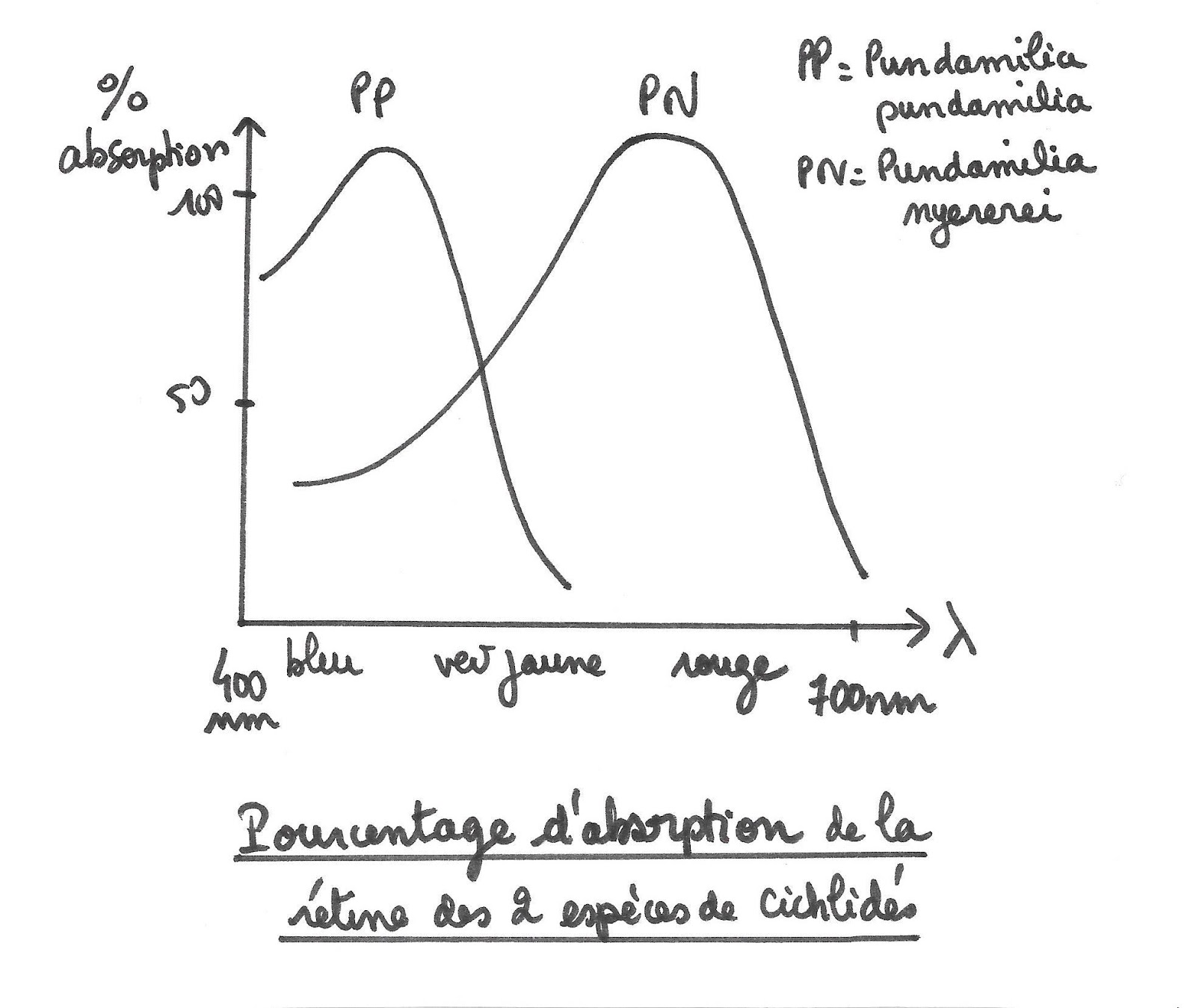
Pourcentages d’absorption de la rétine des deux cichlidés étudiés
L’absence d’hybrides dans le lac montre que Pundamilia nyererei et Pundamilia pundamilia ne se reproduisent pas entre eux, donc ils ne sont pas interféconds, donc ils n’appartiennent pas à la même espèce.
En lumière naturelle, les femelles de P pundamilia choisissent préférentiellement les mâles de couleur bleue et celles de P nyerei choisissent les mâles de couleur vive alors qu’en lumière orange où la distinction des couleurs est impossible, le choix se fait sans discrimination. On peut en déduire que près de la surface, où la sensibilité à la lumière bleue est favorisée, les femelles ayant un spectre d’absorption maximal dans le bleu, peuvent mieux distinguer cette couleur. Les femelles P pundamilia y seront alors prédominantes. De plus les femelles, étant plus sensibles aux radiations bleues que rouges, sont attirées par les mâles ayant une coloration intense. Les poissons rouges apparaissant ternes pour P pundamilia, la reproduction des mâles bleus est donc favorisée. Inversement en profondeur pour les mâles rouges où P nyerei est prédominant.
Ainsi sur de nombreuses générations de sélection sexuelle agissant de cette manière, les deux populations peuvent diverger. Or nous savons que des variabilités des individus pour un certain nombre de caractères entraînent un isolement reproducteur d’une population au sein même de la population mère. Ainsi, bien qu’ils cohabitent dans le même lac, les poissons femelles préféreront s’accoupler en fonction de la couleur des mâles qui les attirent, vu leur sensibilité à la lumière et leur habitat. Au fil des années, les deux sous populations vont cesser de s’accoupler entre elles et établir assez de différences les conduisant à être considérées comme des espèces différentes.



