Cours
Fleur et pollinisation
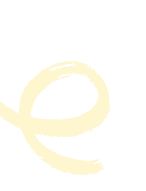



Les plantes à fleurs sont capables de produire par fécondation, des individus génétiquement différents des plantes mères. Cette méthode de reproduction est fortement soumise aux contraintes environnementales. Le processus peut être long et le taux de réussite souvent faible.
La reproduction asexuée est une reproduction rapide qui ne fait pas intervenir de cellules sexuelles.
C’est donc une reproduction obtenue sans fécondation à partir d’un individu parental unique. Les nouveaux individus obtenus sont génétiquement identiques à la plante mère. Ainsi si leur génotype permet un phénotype adapté à l’environnement, ce dernier sera rapidement envahi par un clone de la plante de départ.
La reproduction asexuée ou multiplication végétative au sens large est une multiplication qui s’opère à partir des tissus végétatifs (= non reproducteurs) de la plante parentale et qui peut faire intervenir :
Des organes végétatifs non spécialisés (tiges, racines, feuilles…)
Des organes végétatifs spécialisés : tubercules, stolons, racines drageonnantes…
Le marcottage
Naturel
Le marcottage est une reproduction végétative réalisée par une structure de l’appareil végétatif avant de s’être séparée de la plante mère. Le fragment de végétal impliqué s’appelle une marcotte.
Les rhizomes comme celui du sceau de Salomon, des iris ou du gingembre peuvent réaliser le marcottage naturel même si ce n’est pas leur fonction première.
Un rhizome est une tige souterraine sur laquelle on peut observer des cicatrices foliaires qui sont en fait des feuilles avortées.
Au fur et à mesure de la croissance de ce rhizome des racines adventices se mettent en place préparant ainsi la plante à la fragmentation : si une partie du rhizome se détache, comme elle est pourvue de racines, elle pourra donc survivre et un nouvel organisme sera individualisé. En l’absence de fragmentation, seul le bourgeon terminal produit une pousse feuillée et la plante s’allonge d’une unité chaque année.

Rhizome d’iris montrant un bourgeon terminal développé

Rhizome de gingembre montrant des bourgeons terminaux non développés
Artificiel
Le marcottage artificiel consiste à forcer la mise en contact d'une partie aérienne d'une plante avec un substrat humide, jusqu'à l'apparition de racines. On peut alors « sevrer la marcotte », c'est-à-dire séparer la partie aérienne portant des nouvelles racines de la plante mère. On distingue plusieurs techniques de marcottage :
« par couchage »
Ce marcottage consiste à coucher et maintenir par un arceau au niveau du sol, un rameau souple que l’on aura effeuillé.
Marcottage par couchage
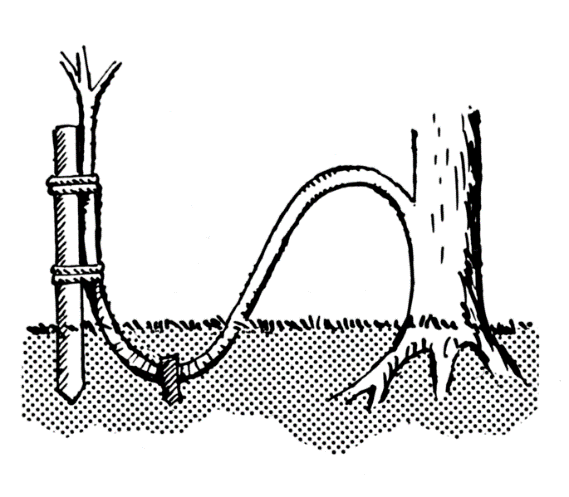
« en butte »
Ce marcottage est adapté aux plantes produisant facilement des rejets. Le processus se réalise sur 2 ans pour les arbustes. Au premier hiver, la plante est rabattue à une dizaine de centimètres du sol ce qui la force à produire des rameaux très proches du sol au printemps suivant. Pendant l’hiver de la deuxième année, une butte de terre est disposée autour de la plante. Cela force l’apparition de racines sur les rameaux enterrés. Ces derniers pourront alors être sevrés et plantés.
Marcottage en butte
« aérien » ou « annulaire »
Ce marcottage est adapté aux arbustes à enracinement difficile et aux plantes d’intérieur. Il consiste à effeuiller le milieu d’une branche, à inciser l’écorce et à disposer tout autour de la zone un manchon de terreau et de sable. En maintenant ce manchon humide, des racines se forment et la branche pourra alors être sevrée. La manipulation fonctionne mieux avec une application d’hormones de bouturage sur les incisions avant l’installation du manchon.

Marcottage aérien
Le bouturage
Naturel
Contrairement au marcottage, c’est une reproduction végétative qui s’effectue à partir d’un fragment d’une plante qui s’enracine après sa chute de la plante mère. Le fragment est appelé une bouture.
On peut citer en exemple le Figuier de barbarie de la famille des cactus et dont les rameaux charnus appelés « raquettes » sont capables de s’enraciner une fois tombés au sol. De ce fait, il devient vite envahissant.

Figuier de barbarie à croissance envahissante

Développement d’un nouvel individu à partir d’une feuille charnue de plante grasse
Artificiel
Il est aisé d’obtenir des boutures chez certaines espèces.
Chez les cactées comme ce cactus du genre Schlumbergera ou cactus de noël, il suffit de planter un rameau dans du terreau pour voir un petit système racinaire apparaître en quelques semaines.

Cactus de noël obtenu par bouturage artificiel
D’autres espèces sont susceptibles de développer des racines au contact de l’eau ou d’un mélange de terreau.
Chez le saintpaulia par exemple, il suffit de sectionner proprement une feuille à sa base et de la repiquer soit dans de l’eau soit dans du terreau. En quelques semaines vous verrez des racines apparaître.

Bouturage artificiel d’une feuille de saintpaulia montrant les racines sur le pétiole de la feuille
La stolonisation
Les stolons sont des organes spécialisés dans la multiplication végétative et c’est là leur unique fonction.
Le stolon présente une croissance plagiotrope, c'est-à-dire perpendiculaire au champ de pesanteur. Son bourgeon terminal donne un nouvel individu à distance de l’individu d’origine. Cette tige est généralement grêle à feuilles réduites et à entrenœuds longs. Quand le stolon entre en contact prolongé avec un sol humide, des racines s’y développent formant un nouveau plant. Une fois le plant enraciné, le stolon s’assèche et la nouvelle plante s’individualise.
On peut donner comme exemples de plantes stolonifères, le fraisier, le lierre et une plante verte bien connue des jardineries, le chlorophytum.

Stolons de fraisier

Stolons de chlorophytum
Les drageons
Les drageons sont des tiges à croissance verticale formées à partir de bourgeons de racines à croissance plagiotrope.
Ces bourgeons sont formés à partir des cellules du péricycle, couche de cellules situées sous l’endoderme et sont à l’origine d’un nouveau système caulinaire et de racines qualifiées d’adventives. Si les drageons sont séparés de la souche mère, ils constituent une nouvelle population de pieds. Ainsi par la production de drageons, la plante mère envahit progressivement l’espace situé autour d’elle.
On peut citer en exemple le framboisier, le peuplier ou encore le sorbier.

Racine plagiotrope de framboisier portant de futurs drageons

Drageons émergeant le long des racines superficielles d’un peuplier après une période de fauche tardive
Les organes tubérisés
Les organes tubérisés ou tubercules sont des organes renflés accumulant des réserves organiques.
Certains d’entre eux peuvent participer à la multiplication végétative.
Les tubercules caulinaires
Les tubercules caulinaires souterrains comme la pomme de terre, présentent des cicatrices foliaires pourvues de bourgeons. Les tubercules caulinaires se trouvant systématiquement à l’apex de tiges souterraines ou de stolons, ils présentent un bourgeon apical à partir duquel se développera une plante feuillée.

Tubercule de pomme de terre germé
Tubercules racinaires
Les tubercules racinaires comme la carotte, hypocotylaires comme le radis ou mixtes participent rarement à la multiplication végétative. Certains le peuvent comme le tubercule racinaire de dahlia ou le tubercule de patate douce. Ce dernier possède dans sa partie proximale raccordée à la racine, de nombreux bourgeons à l’origine de futures pousses.

Tubercule de patate douce
Les bulbilles souterraines
Les bulbes sont des organes tubérisés de nature foliaire qui n’interviennent pas dans la reproduction asexuée mais permettent le passage de l’hiver et un cycle de vie bisannuel.
C’est le cas de l’oignon.
Certains bulbes présentent des unités bulbeuses multiples possédant chacune un bourgeon terminal qui sont en réalité des bourgeons axillaires à l’échelle de tout le bulbe : ce sont des bulbilles. C’est le cas de l’ail. Chaque bulbille est à l’origine au printemps suivant d’un nouvel individu.

Coupe d’une tête d’ail montrant les bulbilles
Les bulbilles aériennes
Des plantes peuvent développer des bulbilles non pas à partir d’une tige souterraine comme pour l’ail, mais à l’aisselle des feuilles chlorophylliennes au niveau du bourgeon axillaire, sur le bord des feuilles ou au niveau de fleurs transformées en bourgeons adventifs, mais dans ce cas ces bourgeons adventifs n’ont pas de réserves.
Les bulbilles se détachent de la plante mère à maturité et produisent un nouvel individu.

Les bulbilles aériennes foliaires du kalanchoé
L’édification d’un nouvel individu implique le fonctionnement de zones méristématiques où des cellules indifférenciées subissent de nombreuses mitoses, à l’origine des cellules du nouvel individu, puis une différenciation, assurant la mise en place des différents tissus et organes du nouvel individu.
Quand la formation des tissus du nouvel individu se fait à partir de tissus déjà différenciés constitués de cellules moyennement différenciées (= encore vivantes, sans lignine, sans épaississements cellulosiques excessifs…), ces dernières vont subir un retour à l’état méristématique : c’est la dédifférenciation.
Il y alors néoformation de méristèmes. Différents signaux d’origine interne (phytohormones) et/ou externes peuvent induire une reprogrammation génétique de la cellule, entraînant la perte des caractères différenciés puis d’autres signaux entraînent la différenciation des nouvelles cellules.
En laboratoire, pour former un nouvel individu à partir d’un fragment végétal, il faut donc d’abord obtenir une masse de cellules indifférenciées appelée « cal». C’est à partir de ce cal que les techniciens de laboratoire horticole vont provoquer soit la rhizogenèse (formation de racines) soit la caulogenèse (formation de tiges). Pour cela, ils vont utiliser les hormones de développement comme les auxines et les cytokinines. Les concentrations relatives de ces 2 hormones déterminent la différenciation des cellules du cal.
La callogenèse est activée quand le rapport AIA/ CK est = 1.
La rhizogenèse est activée quand le rapport AIA/ CK est >1, bloquée quand elle est <1.
La caulogenèse est activée quand le rapport AIA/ CK est <1, bloquée quand elle est >1.
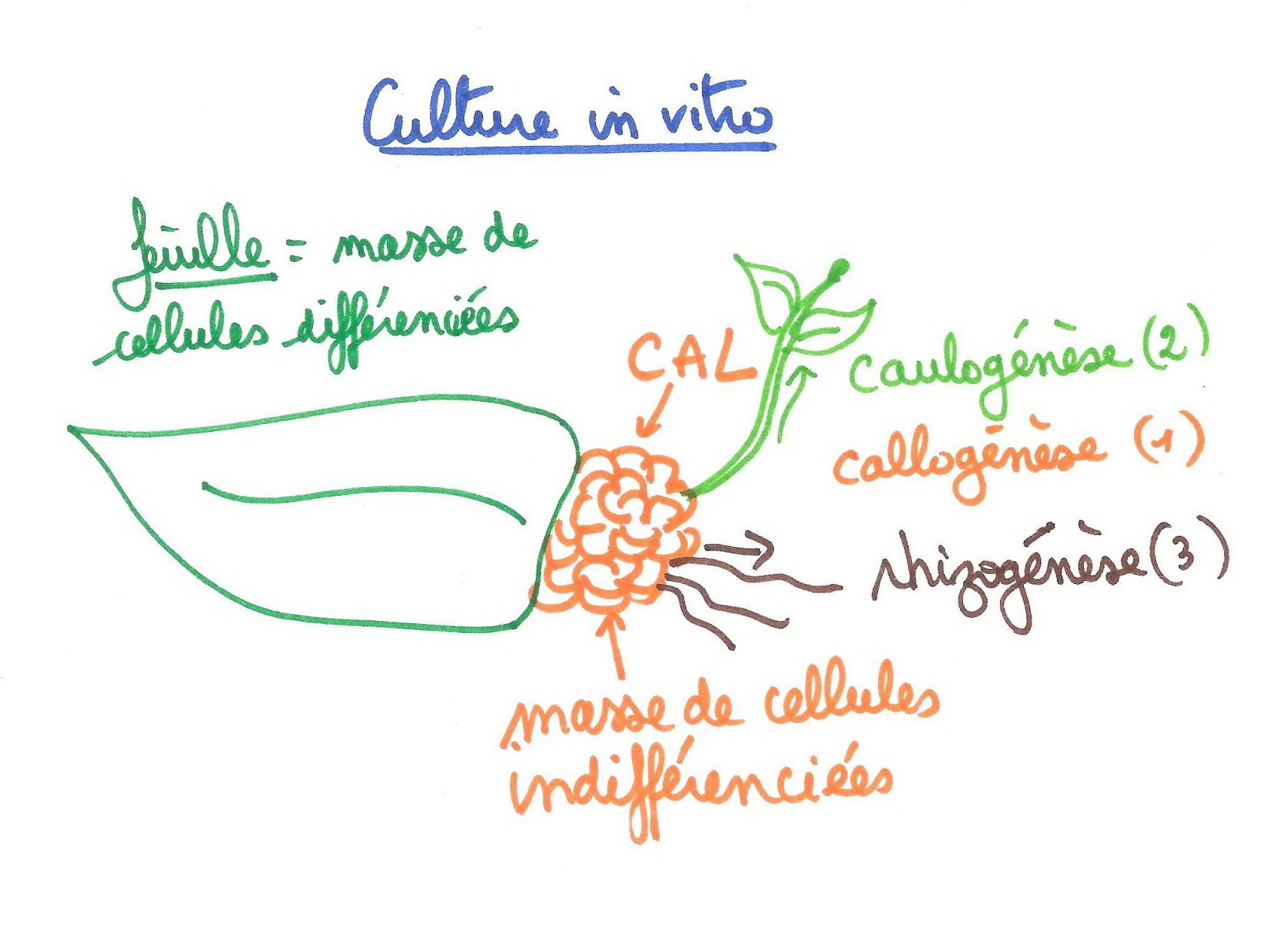
Principe de la culture in vitro
Il est ainsi possible en milieu de culture stérile de forcer l’apparition de méristèmes à partir de fragments de végétaux en appliquant les bons dosages hormonaux.

Culture in vitro de plant de patates douces



