Cours
Les lois de Mendel




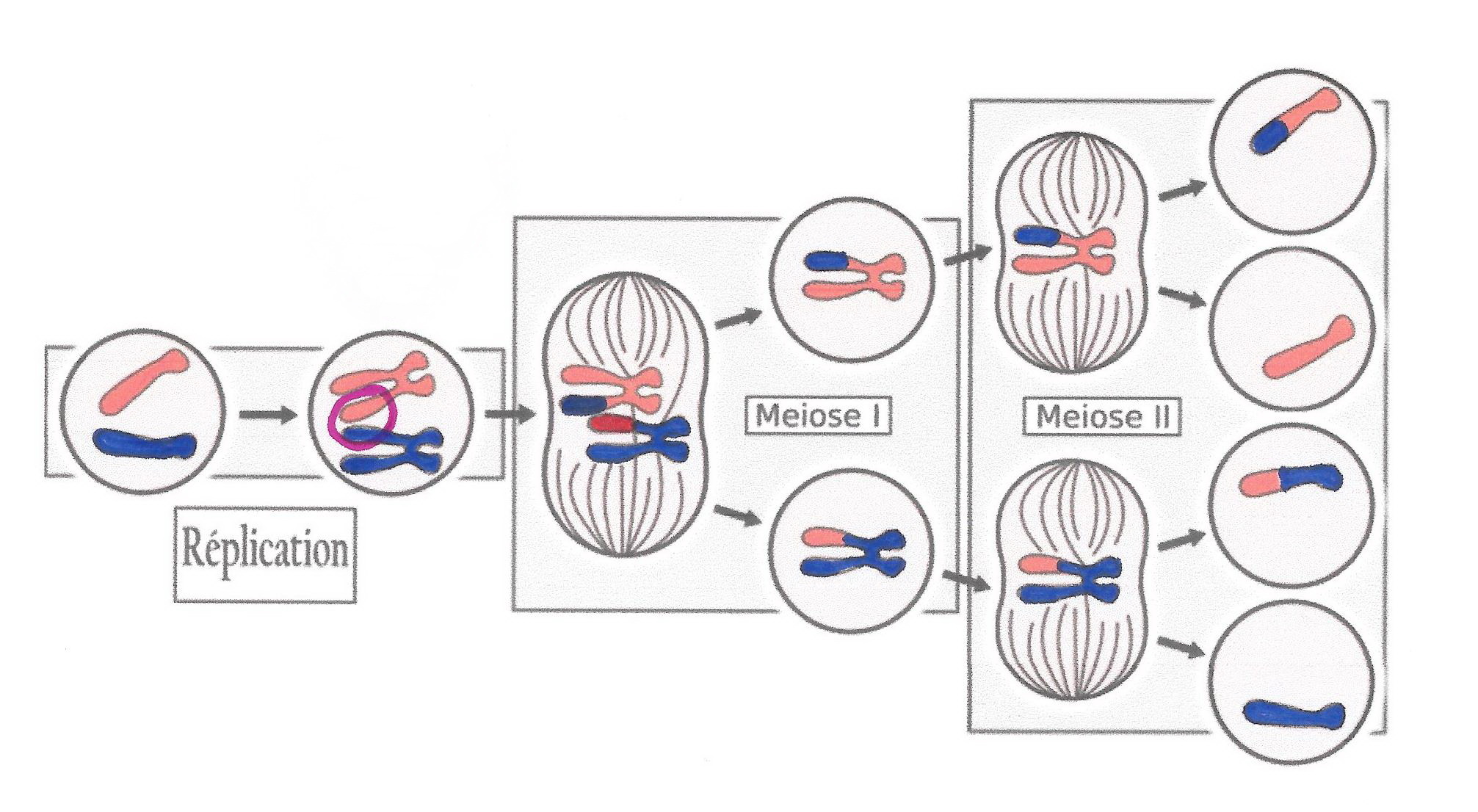
L’étude du graphique montrant l’évolution de la quantité d’ADN dans une cellule germinale allant subir une méiose révèle, après un épisode de duplication, deux divisions successives.
La première division est une division dite « réductionnelle » car elle consiste à séparer les deux exemplaires de chaque paire de chromosomes doubles.
La deuxième division est une division dite « équationnelle » car la fracturation des chromosomes doubles en chromosomes simples et leur distribution à chacune des 2 cellules filles, permet de conserver un exemplaire de chaque sorte dans chacune d’elle.
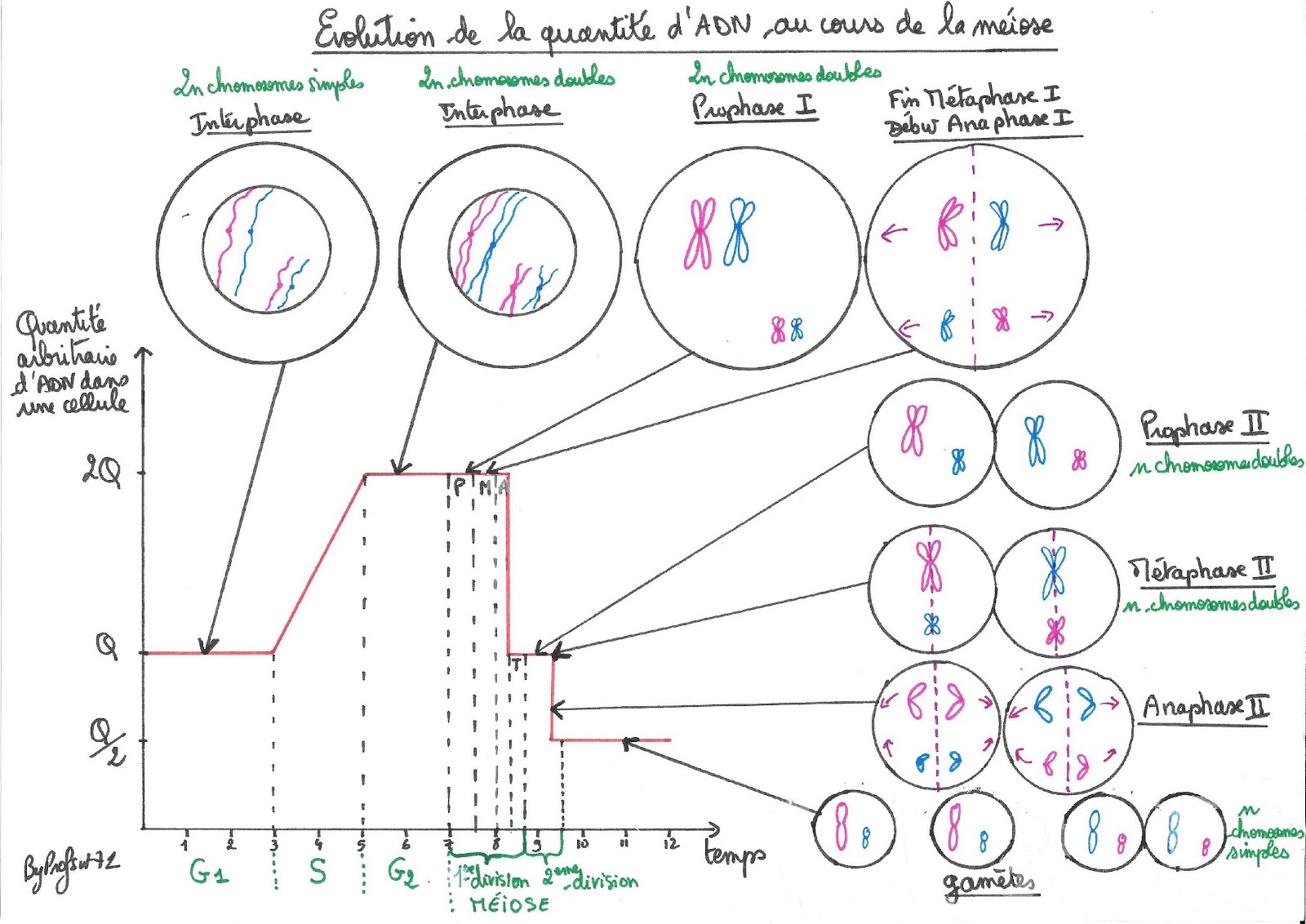
Évolution de la quantité d’ADN au cours de la méiose
La méiose est donc une division cellulaire qui permet le passage de la diploïde à l’haploïdie, c’est-à-dire d’une cellule qui possède deux lots de chromosomes doubles à 4 cellules qui ne possèdent qu’un lot de chromosomes simples.
Ces cellules possédant « n » chromosomes simples sont des cellules reproductrices ou gamètes chez la plupart des eucaryotes. La fécondation réunira ainsi deux lots de chromosomes et la cellule-œuf obtenue aura donc un caryotype à 2n chromosomes.
Chez l’Homme, la méiose ayant lieu uniquement dans les cellules germinales des testicules ou des ovaires il est impossible de suivre en direct le brassage génétique lié à la distribution des 23 paires de chromosomes.
C’est en revanche plus facile de le faire chez certains organismes simples comme le champignon Sordaria macrospora.
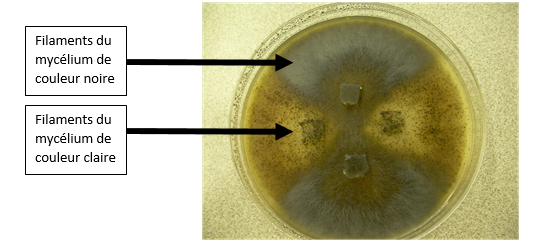
Culture de Sordaria claire et de Sordaria noire sur une même boite de pétri.
Ce champignon de l’embranchement des ascomycètes, présente la particularité d’avoir un cycle de vie majoritairement haploïde. La germination d’une spore engendre un filament mycélien : les cellules se forment par mitoses à partir de la cellule contenue dans la spore. Ces cellules sont haploïdes.
Quand deux filaments se rencontrent, il y a fusion des cellules entrées en contact suivie d’une caryogamie (fusion des noyaux) ce qui correspond au final à une fécondation.
Cycle de vie de Sordaria macrospora
La cellule formée correspond en quelque sorte à une cellule-œuf à partir de laquelle va se former une structure appelée un « périthèce » dans lequel vont se développer des asques, sortes de manchons protecteurs contenant chacun huit spores. En installant un périthèce entre lame et lamelle et en exerçant une faible pression sur cette dernière, celui-ci va s’ouvrir libérant les asques qu’il contient.
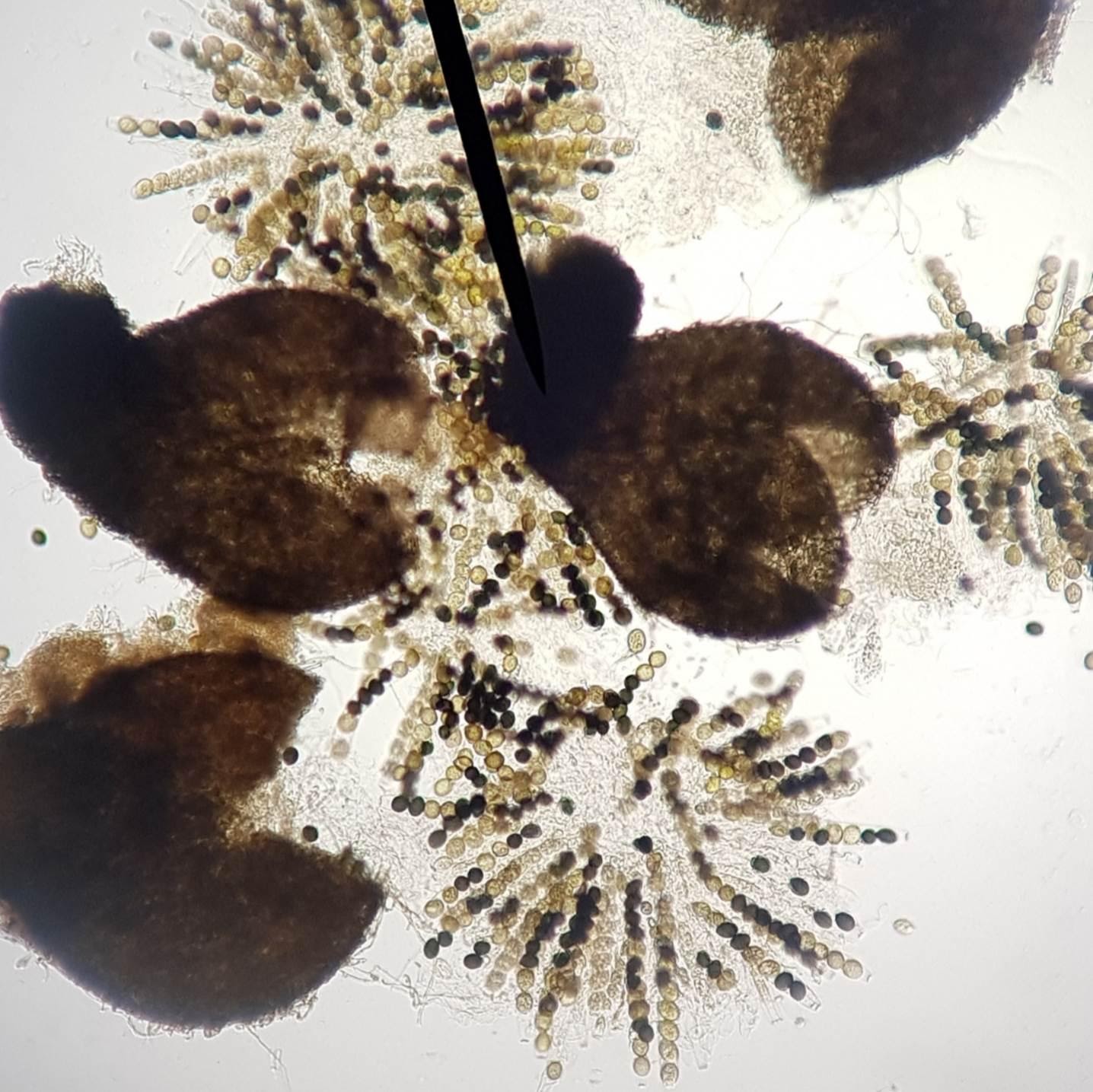
Périthèces ouverts de Sordaria macrospora et asques contenant 8 spores chacunes libérées
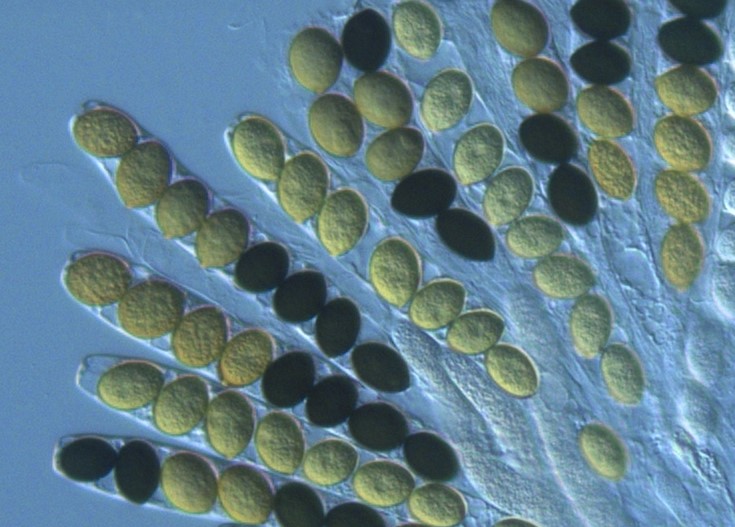
Asques de Sordaria macrospora
Ces spores sont constituées d’une cellule haploïde ce qui signifie qu’elles ont été formées par méiose. Cependant une méiose ne produit que 4 cellules or ici il y en a 8, il y a donc eu une méiose suivie d’une mitose. Ces spores peuvent être noires ou claires et ce caractère est contrôlé par un gène existant en deux versions : l’allèle noir (n) et l’allèle clair (c). Du fait que les spores soient immobilisées dans l’asque, il est possible en étudiant leur couleur et leur position de retrouver le génotype de la cellule de départ ainsi que les modalités de distribution des chromosomes lors de la méiose.
Travaillons sur des asques obtenus à partir d’une cellule diploïde issue de la fusion d’une souche de Sordaria à spores noires et d’une souche de Sordaria à spores claires.
Prenons le cas d’un asque constitué à sa base de 4 spores noires surmontées par 4 spores claires (doc ci-dessous). Il y a 2 couleurs de spores ce qui signifie que la cellule de départ possédait les 2 allèles n et c : elle était donc hétérozygote. Si l’asque n’avait contenu que des spores noires cela aurait signifié que la cellule de départ était homozygote pour l’allèle noir. Idem pour 8 spores claires.
La position des spores indique que lors de la première division de méiose, le chromosome portant l’allèle clair a été distribué dans la cellule du haut et le chromosome portant l’allèle noir a été distribué dans la cellule du bas. La deuxième division n’a fait que séparer les chromatides identiques de chaque chromosome double donnant ainsi deux spores voisines de même couleur car exprimant le même allèle. La mitose suivante n’a fait que doubler les cellules.
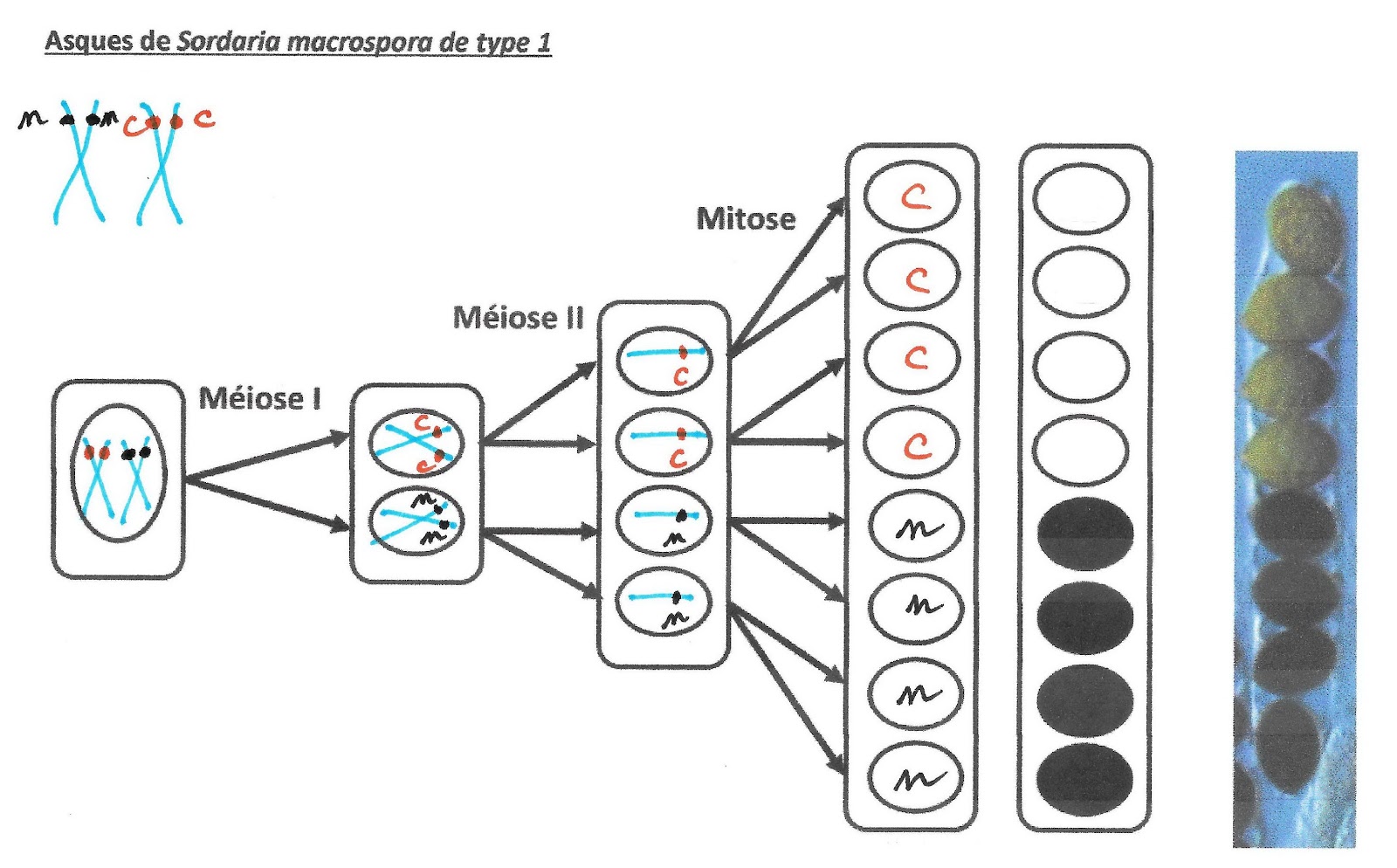
Asques de Sordaria macrospora de type 1
Pour l’exemple n°2 ci après, la cellule de départ est aussi hétérozygote mais la distribution de chromosomes doubles a été inversée : le chromosome portant l’allèle noir a cette fois été distribué dans la cellule du haut.
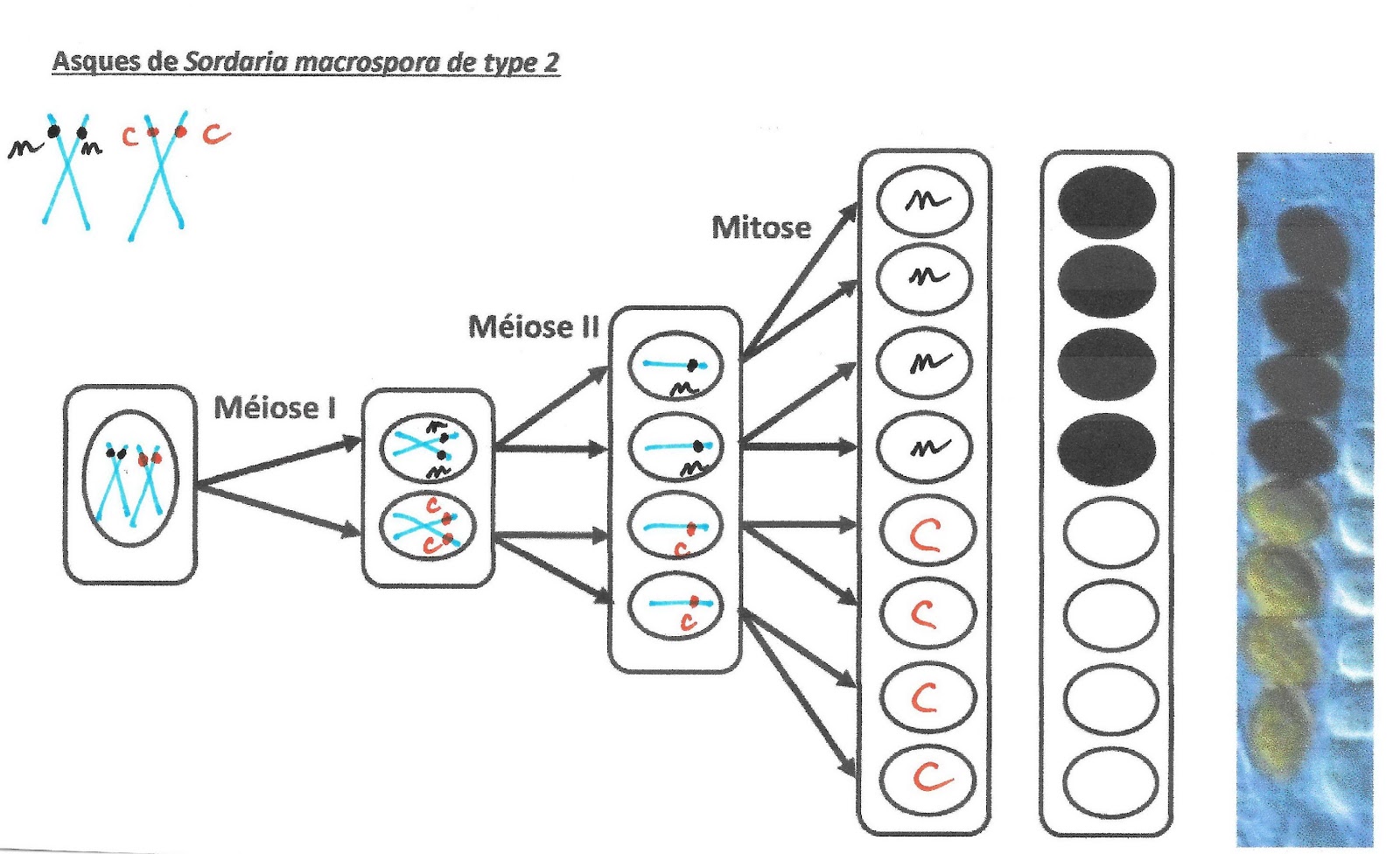
Asques de Sordaria macrospora de type 2
Il y a donc pour une cellule hétérozygote, 2 possibilités équiprobables de distribution des chromosomes homologues de part et d’autre de la plaque équatoriale de la cellule-œuf de départ. On parle de brassage inter-chromosomique car le brassage des informations, se joue au niveau de la distribution des chromosomes de chaque paire.
Il existe un brassage dit intra-chromosomique qui se déroule au sein d’une paire de chromosomes.
Les chromosomes homologues échangent des portions de chromatides : ils échangent donc des allèles. Ainsi, si une paire de chromosomes présente une hétérozygotie pour un gène, les conséquences de cet échange de matériel (appelé aussi crossing-over), créé des chromosomes doubles portant des allèles différents sur le même locus alors que normalement après réplication ce n’est pas le cas.
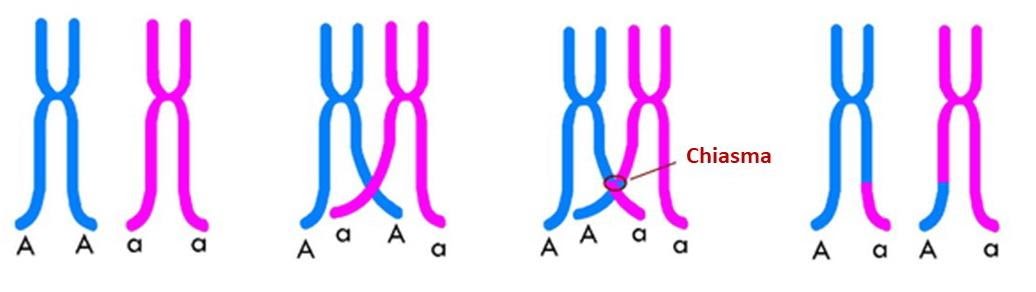
Mécanisme du crossing over
Ce mécanisme va provoquer une variation de la disposition des spores de couleur dans l’asque. Leur position permet de retrouver comment s’est déroulé le crossing-over puis la distribution des chromosomes et des chromatides comme il est possible de le voir sur les 4 exemples suivants.
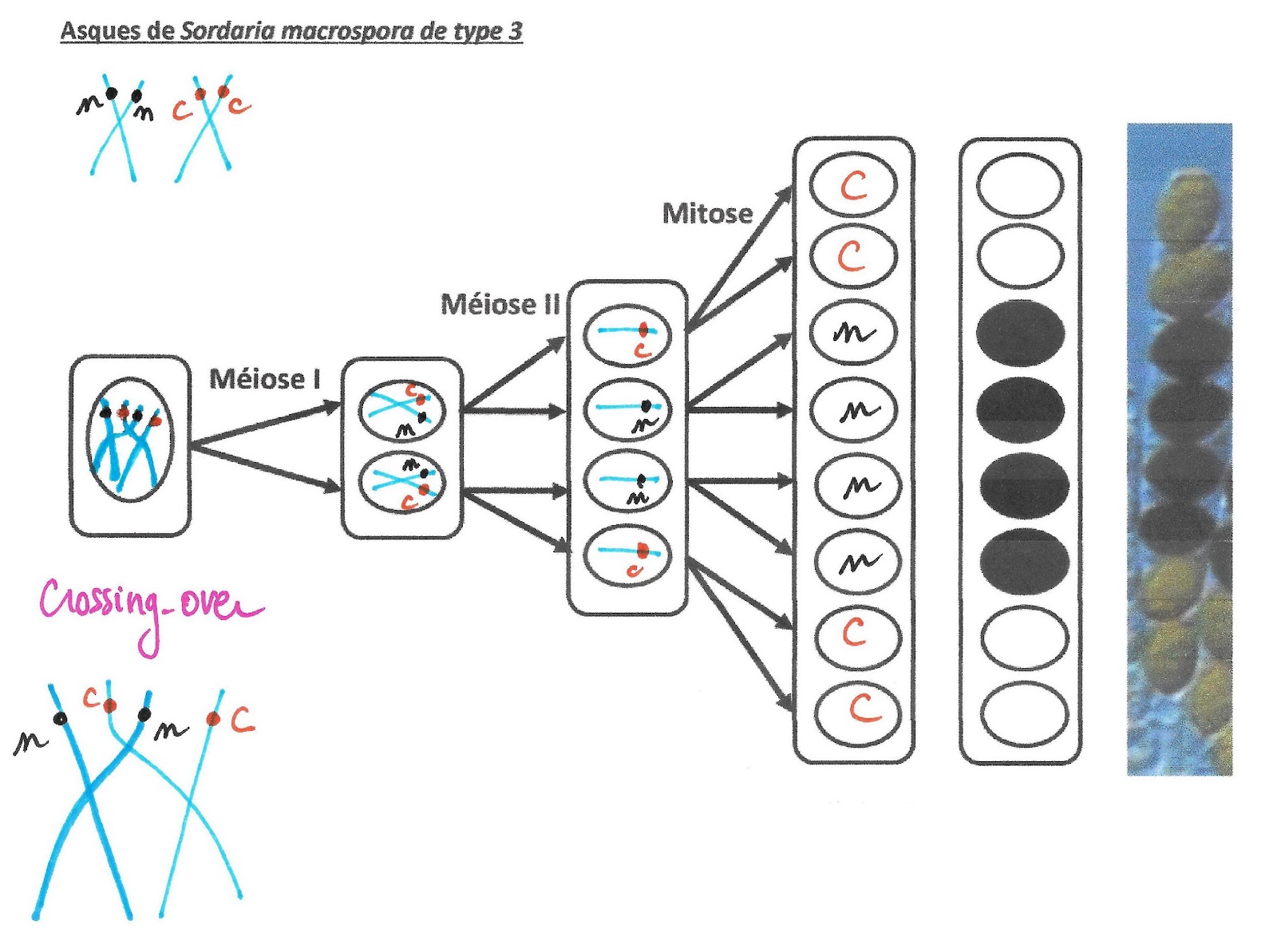
Asques de Sordaria macrospora de type 3
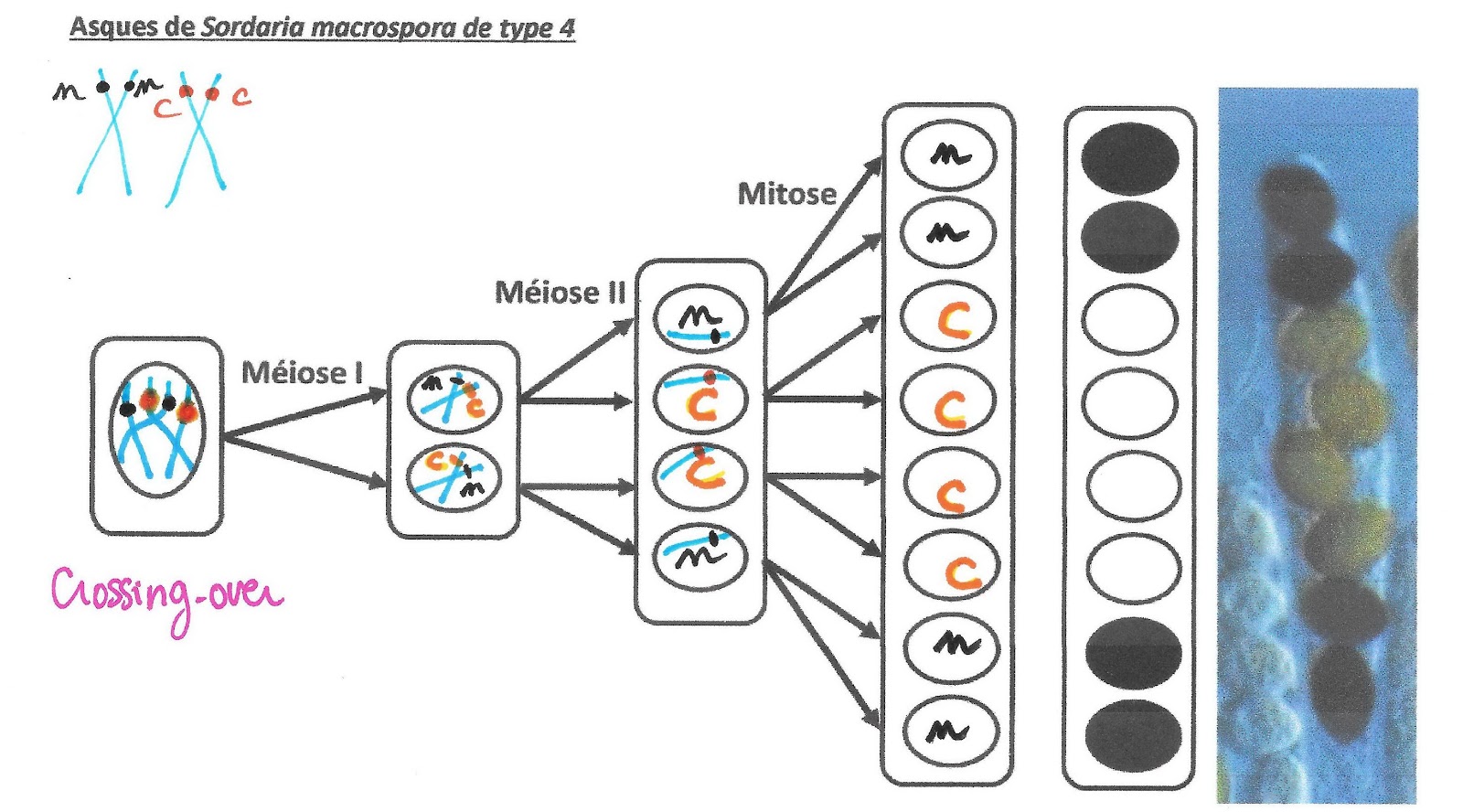
Asques de Sordaria macrospora de type 4
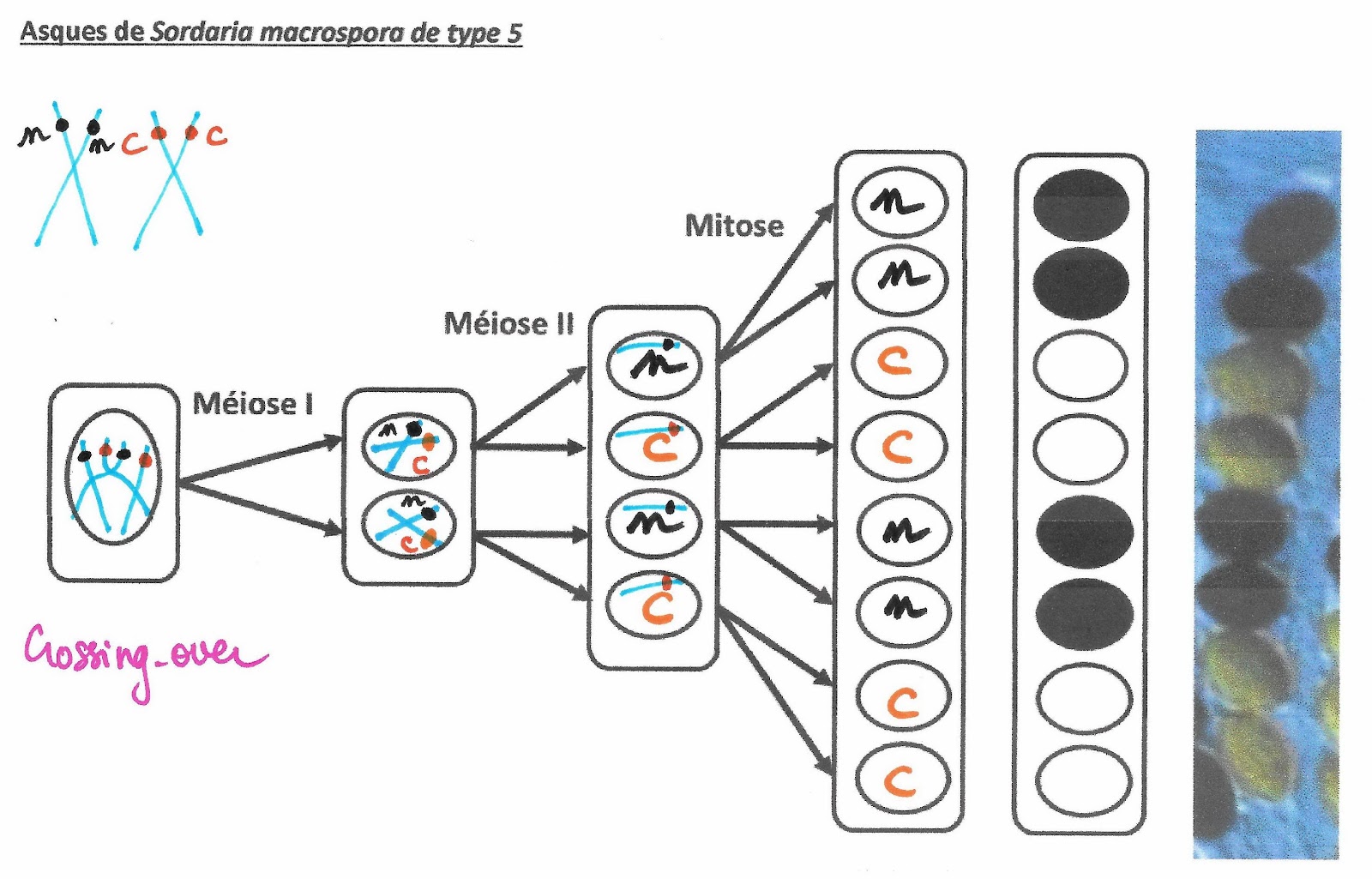
Asques de Sordaria macrospora de type 5
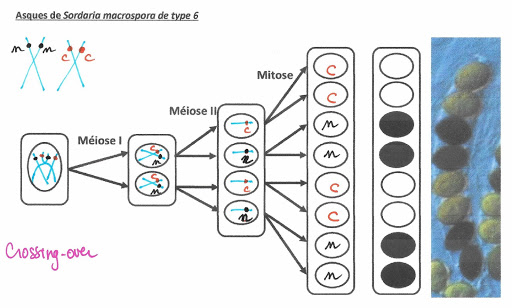
Asques de Sordaria macrospora de type 6
Si l’on revient aux êtres vivants dont la majeure partie du cycle est diploïde, comme chez l’Homme, le brassage inter-chromosomique est systématique dans les cellules germinales en méiose. Le nombre de combinaisons possibles est d’autant plus élevé que le nombre de chromosomes et le nombre de gènes à l’état hétérozygote est plus grand chez les parents.
Dans 10 à 12% des cas, au début de la méiose, à la prophase 1 (prophase de la première division de méiose), il y a un brassage intra-chromosomique. C’est la combinaison des deux brassages qui fait la diversité des êtres vivants.
Méiose et brassages chez l’Homme
Sans tenir compte du brassage intra-chromosomique, il est possible de calculer le nombre de spermatozoïdes ou d’ovules différents qu’un homme ou une femme peut produire ainsi que le nombre théorique d’enfants génétiquement différents qu’un couple peut produire.
En tenant compte du fait que nous possédions 23 paires de chromosomes et qu’au moment de la méiose chaque paire présente deux possibilités de distribution, il y a alors
Lors de la fécondation,
gamète de la mère a la possibilité de rencontrer
gamètes différents du père.
Donc les 2^23 gamètes de la mère peuvent rencontrer chacun 2^23 gamètes différents du père donc le nombre de fécondations possibles correspond à une combinaison soit:

Arbre de distribution pour la formation des gamètes pour 2 ou 3 paires de chromosomes



